Hammerhead ribozyme motif is made up of three base-paired helices (I-III) and a core of highly conserved, non-complementary nucleotides necessary for the catalysis. The hammerhead ribozyme catalytic motif was discovered by comparing the sequences of certain plant viroids, which undergo site-specific, self-catalyzed cleavage as part of their replication process (Birikh, 1997). The secondary structure resembles the shape of a hammerhead (Fig.1) and serves as its distinct trademark. As discussed earlier, the 'single turnover' ribozyme has to be genetically engineered to perform 'multiple turnover' by separating a substrate strand, which contains the cleavage site, from the catalytic strand.
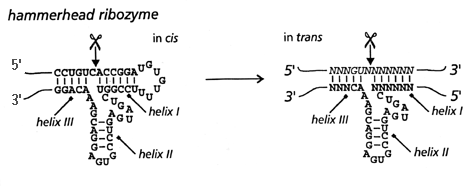
Figure 1. Hammerhead ribozyme (in cis) and its engineered version (in trans)
The secondary structure of a ribozyme can be predicted from the primary sequence by analyzing the base-pairing and free energy reduction during the folding. Examples of available software are RNAdraw and mfold. The sequence of the in cis ribozyme in Fig.1 was manually input to mfold and Fig.2 shows how the computed graphic looks. Also, the BLAST search was performed and see the results from here.
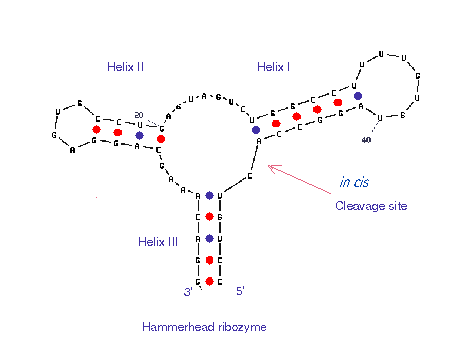
Figure 2. A secondary structure calculated by mfold. The structure is identical with the in cis ribozyme shown in Fig.1, though the orientation looks different. Also see the corresponding loop free energy decomposition.
X-ray crystallography has been used successfully in solving some hammerhead ribozyme 3-dimensional structures. A primary problem is: how to maintain stable enzyme-substrate hybrids in the crystal without having the substrate strands cleaved? Two general methods were applied in achieving this, (1) by using DNA (Due to the absence of 2'-OH group in DNA, the catalytic intermediate can not be formed) as the substrate to get a RNA/DNA hybrid (Pley, 1994, Fig.3) or (2) by changing [Mg2+] and pH to eliminate the enzymatic activity (Scott, 1995 Fig.4).
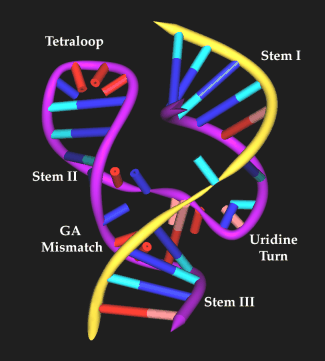
Figure 3. The hammerhead ribozyme (RNA/DNA) structure by Pley, Flaherty and Mckay(1994), purple ribbon shows the RNA ribozyme where as the yellow ribbon represents DNA strand.
The conserved catalytic core contains two RNA structural motifs. The first involves nucleotides C3 to A6 which form a 'uridine turn' or 'U turn'. This U-turn conformation is stabilized by several hydrogen-bonding interaction (U4(2-OH) -A6 (N7), U4 (N3)-U7(P)). The second motif in the catalytic core is a duplex containing tandem GA mismatched base pairs (G8-A13, A9-G12). The large helix and twist angle between the two GA base pairs lead to stacking of G8 on G12 and stacking of A9 on A13. The crystal structure of the hammerhead also revealed that the GA duplex domain has a divalent metal-binding site. It is widely accepted that mismatched region are often highly structured and can readily form non-Watson-Crick base pairs. The hammerhead ribozyme contains another RNA folding motif, a GAAA tetraloop. It is not a consensus structure but was designed to form a stable structure at the end of helix II. Additional studies have shown that GNRA tetraloop may act as protein binding site and are involved in tertiary interaction. GAAA tetraloop can also form an intermolecular interaction with CG base pairs on stem II(as shown in Fig. 3) (Pardi, 1994).
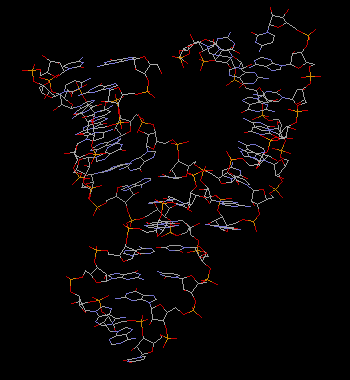
Figure 3. Hammerhead ribozyme structure (RNA/RNA) by Scott et al (1996). Yellow lines show the backbone of the two nucleotide chains (from Rasmol).

Figure 4. RNA Hammerhead Ribozyme (Tuschl,1994),
made by Microsoft GIF Animator.