






|
|
|
|
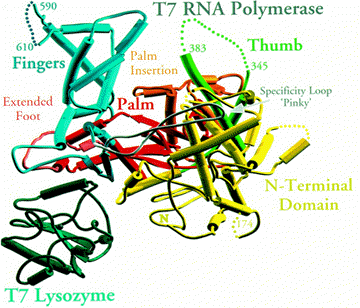
Overall Architecture: The structural characterization of bacteriophage T7 RNA polymerase has been elucidated through biophysical, biochemical and genetic studies. Four separate crystal structures have been solved for the enzyme: the uncomplexed enzyme (Sousa et. al, 1993), the enzyme complexed with its inhibitor T7 lysozyme (Jeruzalmi et. al., 1998), a binary complex of the polymerase bound to its promoter(Cheetham. al., 1999a), and an initiation complex in which the first three nucleotides of RNA have been synthesized (Cheetham et. al., 1998b). The crystal structure of the polymerase alone was determined at 3.3 � resolution by Sousa et. al., 1993. The single-subunit enzyme is highly a-helical (Figure 4a). It is organized around a cleft with the dimension to accommodate a double-stranded DNA template (Figure 4, Figure 6, Figure 7, and Figure 8). The enzyme consists of a polymerase and an N-terminal domain (Figure 5A). The polymerase domain has a U-shape fold, a feature that is characteristic of all pol I polymerases (Kohlstaedt, 1992). Similar to other nucleic acid polymerases, the RNA polymerase domain resembles an open right hand and, as a result, it has further been divided into a palm, thumb, and a fingers domain. The palm, fingers and thumb domains of the T7 RNA polymerase define the DNA binding and RNA synthesis catalytic site [View Chime Image] [see Note].
Figure 4. The structure of T7 RNA polymerase alone. In (a), the long axis of the active site is parallel to the plane of the image, while in (b) it is perpendicular to the plane of the picture (Sousa et. al, 1993).
The thumb domain comprises amino acids 325-411 (Figure 5B). It forms a primarily a-helical extension from the palm domain on one side of the template binding cleft. This domain is very flexible, and it has been demonstrated that it functions to stabilize the ternary complex during processive transcription by wrapping around bound template. Mutant T7 RNA polymerases with shorter thumbs have been shown to be less processive, presumably due to a lower affinitity for the template DNA (Bonner et. al, 1994).
Figure 5. Schematic representation indicating the structural domains of T7 RNA polymerase. The palm domain consists of amino acid residues 412-565 and 785-883 (Figure 5B). This domain is situated at the base of a deep cleft that is bounded by the fingers and the thumb domains (Figure 4a, Figure 6, and Figure 7). It contains a trio of b-strands that are conserved in every nucleic acid polymerase (Steitz, 1994). These b-strands contains the catalytically most critical and conserved amino acids, a pair of aspartate residues, Asp537 and Asp812, which are directly involved in catalysis via the interaction with two Mg(II) ions. These two aspartate amino acids are shown in Figure 4a as a red sphere and as a dark blue sphere, respectively. Consistent with the importance of these apartate residues, catalysis of the mutant T7 RNA polymerase deficient in Asp537 and and Asp812 is severely affected without compromising the affinity for rNTPs (Bonner et. al., 1992). It has been reported that the palm domain also plays an important role in sugar discrimination (Gao et. al., 1997).
Figure 6. Structure of T7 RNA polymerase-T7 lysozyme complex (Jeruzalmi et. al., 1998). The fingers domain includes amino acid residues 566-784. This domain raises above the palm domain forming a wall on the left of the active-site cleft (Figure 6, Figure 7, and Figure 8). Relative to the Klenow fragment polymerase, the fingers domain of T7 RNA polymerase contains some insertions. One of these insertions is the 'pinky' specificity loop which is involved in direct base-specific interaction with the major groove of the promoter DNA (Figure 7 and Figure 8). The fingers domain also harbors important catalytic residues such as Tyr639 and Lys631 that are in the proximity of the active site. The relative positions of Tyr639 and Lys631 are indicated in the crystal structure determined by Sousa et. al. as a pink and a light blue spheres, respectively (Figure 4a).
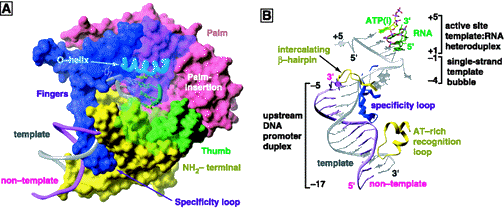
Figure 7. Surface representation of the binary complex of T7 RNA polymerase bound to its promoter DNA (Modified from Cheetham et. al., Nature (1999): 399, 80-83.
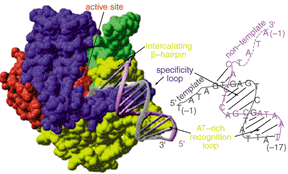
The N-terminal domain of T7 RNA polymerase encompasses amino acid residues 1-324. This domain is positioned in front of the polymerase domain and forms the front wall of the catalytic cleft. It gives the enzyme its characteristic concave shape (Figure 4a and Figure 6). Biochemical, mutational, and x-ray crystallographic analyses have established a role of the N-terminal domain in interaction with upstream regions of promoter DNA and the nascent RNA transcript (Muller et. al., 1988; Cheetham et. al., 999a and 1999b). These studies have demonstrated the involvement of this domain in promoter recognition and DNA melting via the participation of a AT-rich recognition loop and an intercalating b-hairpin loop (Figure 7, Figure 8, and Figure 9).
Figure 8. In (A), surface representation of the T7 RNA polymerase initiation complex; in (B), illustration of how important motifs of the polymerase interact with the promoter DNA (Cheetham et. al., 1999b).
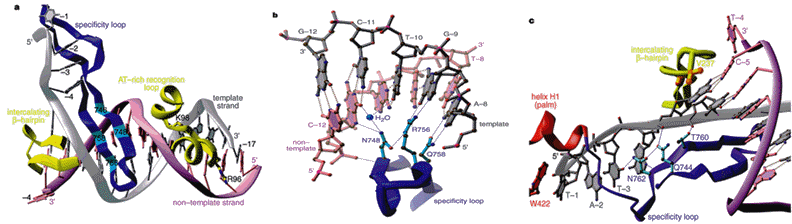
The specificy loop, AT-rich recognition loop and the intercalating b-hairpin loop are three motifs that are unique for the T7 RNA polymerase. They endow the polymerase with a stringent specificity for its own promoter. The result is that, in the presence of Mg(II) ions and rNTPs, the polymerase is self-sufficient to recognize its promoter and synthesize RNA in the absence of any protein factor. The specificy loop is an anti-parallel b-ribbon structure which comprises amino acid residues 739-770. This loop projects into the DNA binding cleft of the T7 RNA polymerase and makes direct base-specific interactions with the upstream region of the promoter during binding and initiation. It specifically inserts into the DNA major groove (Figure 7, Figure 8, and Figure 9a). Some of the amino acid residues of the specificity loop involved in base-specific recognition with the promoter DNA include Gln748, Rrg746, Rrg756, and Asn758. The guanidinium group of Arg756 makes two contact interactions with the base pair at position G-9, while Gln758 makes hydrogen bonding interaction with the base pair at A-8 of the template strand. Asp748 directly interacts with the non-template strand at position G-11, and indirectly, through a water-mediated interaction, with the template and non-template strands at T-10 and G-11, respectively (Figure 9).
Figure 9. In (a), interaction of the specificity loop, AT-rich recognition loop and the intercalating b-hairpin loop with the promoter; in (b), base-specific interaction between the specificity loop and the promoter; and in (c), formation of the transcriptional bubble (Cheetham et. al., 1999b; [See Chime Image].
The AT-rich recognition loop, which comprises residues 93-10, indirectly binds through its inherent flexibility to the DNA sequence in the minor groove at base pair -17 and -13. As depicted in Figure 9a, the side chains of Arg96 and Lys98 interact with phosphate groups causing a distortion to the phosphodiester backbone of the minor groove (Cheetham et. al., 1999b). The intercalating b-hairpin loop plays an important role in transcription initiation: formation and stabilization of the transcription bubble. It facilitates melting of the promoter duplex so that the template strand can be accessible to the binding site of the T7 RNA polymerase. The denaturation of the duplex double stranded DNA is accomplished by the insertion of the b-hairpin loop which harbors the amino acid Val237. Val237 acts directly to separate the template and non-template strands and to position the exposed template towards the active site. As indicated in Figure 9c, the b-hairpin loop intercalates at base pair -4, while the side chain of Val237 makes stacking interaction with base pair -5. The transcription bubble, which is confined to base pair -1 to - 4, is further stabilized by interactions between the single stranded template and the specificity loop (Cheetham et. al., 1999b).
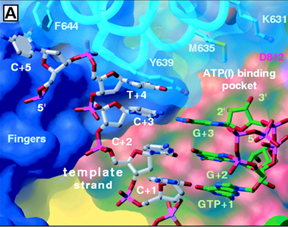
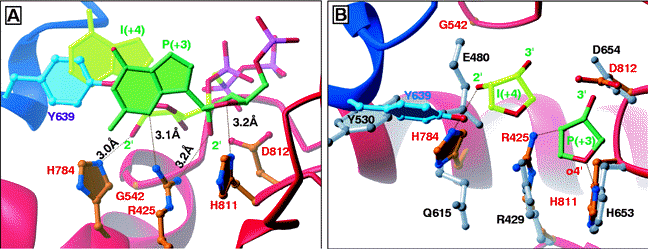
The Active Site: The active site of T7 RNA polymerase is located in a deep pocket that is bounded by the fingers, thumb, palm, and the N-terminal domain (Figure 7 and Figure 8). As indicated above, the active site harbors the catalytically most important residues. It contains two aspatate residues that are involved directly in catalysis; it has Tyr639 which discriminates against sugar by forming hydrogen-bonding interaction with the 2'-OH group of the incoming ribonucleotide. Since deoxyribonucleotides lack a 2'-OH group, they are not incorporated into the groing RNA transcript (Figure 10 and Figure 11). Figure 10. A close view of the active site of T7 RNA polymerase (Cheetham et. al., 1999b). The role of His784 as illustrated in Figure 11 is controversial. Cheetham et. al. (1999b) postulated that this residue also functions as a sugar discriminator by forming hydrogen-bonding interaction with the 2'-OH group of the incoming rNTP. Brieba, et. al. (2000) found, however, that although the His784 point mutant decreases the activity of the polymerase, its effect as a sugar discriminator is not significant. Figure 11. In (A), His784 as a possible sugar discriminator; in (B), comparison between the active sites of T7 RNA polymerase and T7 DNA polymerase (Cheetham et. al., 1999b). |